The aim of this lesson is to give a concise, but detailed overview of how the cell cycle
is regulated. Most importantly, it outlines the roles of the two most important genes that are known to be
key to its regulation: TP53 and Rb. Further, the lesson focuses on the mechanisms implicated in apoptosis –
the process of programmed cell death, whose dysregulation is important in the pathogenesis of cancer.
Some hints and tips on how to approach this topic can be found at the end of the lesson.
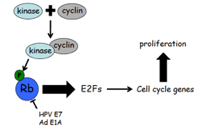
Two main tumour suppressor genes P105-Rb retinoblastoma protein Pocket protein that binds and
inactivates the E2Fs transcription factors that drive the expression of cell cycle genes It provides
the classical model for a recessive tumour suppressor gene in that both paternal and maternal copies of
the gene must be inactivated for the tumour to develop This is called the loss of heterozygosity
(LOH) and can occur in inherited retinoblastoma heterozygous Rb alleles exhibit a high frequency of
LOH Mechanisms for exchange of genetic information between the paired homologous chromosomes in Rb
heterozygote are mitotic recombination, chromosomal non-dysjunction, gene conversion Individuals
with inherited retinoblastoma are also susceptible to malignant tumours of mesenchymal tissues
Inactive Rb alleles are common in small lung cell carcinoma and occur in non-small cell lung, bladder,
breast and pancreatic carcinomas Cell cycle regulation
Rb normally inhibits E2F TFs required for expression of genes in DNA replication P53 tumour
suppressor (TP53 gene) Transcription factor that responds to stress/damage/oncogenic signalling and
induces growth arrest and cell death It is also sometimes called the guardian of the genome as it
ensures that if there is damage to DNA, the cell is arrested and may be diverted into the apoptotic
pathway Mutations in p53 causing loss of function occur in over 70% of human cancers The rare,
autosomal dominant Li-Fraumeni syndrome arises from p53 mutations inherited through the germ line
50% develop diverse cancers by 30 years of age, compared with 1% rate in the general population In
general, p53 mutations are somatic and occur with high frequency in all types of lung cancer, in over
60% of breast tumours and in about 40% of brain tumours (astrocytomas), frequently in combination with
the activation of oncogenes
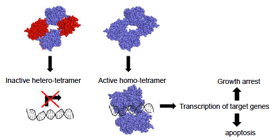
Mechanism of action: Acts as a transcription factor for genes that cause cell cycle arrest in G1 or
those that cause apoptosis
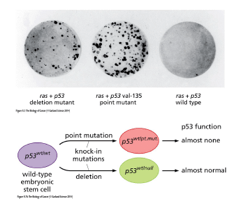
Unlike in Rb, loss of function of p53 is not always dependent on inactivation of both alleles Since
p53 binds DNA mutations in the DNA binding region will be enough to stop it from performing its normal
tumour suppressing action
active p53 binds DNA as a homo-tetramer
The survival plot shows the importance of p53 in cancer outcome In addition to DNA mutations,
epigenetic events can contribute to tumour development For example, tumour cell DNA is generally
hypomethylated (active chromatin conformation) compared with normal cells that are methylated – apart
from hypermethylated regions that are silenced in tumours (often tumour suppressor genes) Importance
of understanding cell proliferation and cell death Cell division is essential for growth of tissues
and organisms -> cells must duplicate their essential components before undergoing cell division to
produce identical daughter cells If this is not controlled properly, tumours may develop ->
uncontrolled proliferation of cells is one feature of cancer cells Therefore, in order to understand
cancer pathogenesis, we need to understand how the cycle is initiated and regulated and which components
are mutated and dysregulated in cancers Many essential regulatory components of the cell cycle prove
to be good targets for therapy Cell death is essential for sculpting of tissues and removal of
damaged cells Cancer is also a disease of resistance to cell death Thus, there needs to be a
balance between cell proliferation and cell death If mitosis proceeded without cell death, an 80yo
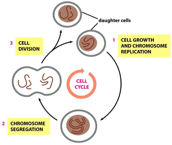
person would have about 2km2 of skin Phases of the cell cycle The purpose of the cell cycle is
to produce two genetically identical daughter cells First, the cell grows and the genetic
information replicates via DNA replication mechanisms Then, chromosome segregate whilst they are
preparing for cell division The last step is cell division into two daughter cells that both share
the same genetic information
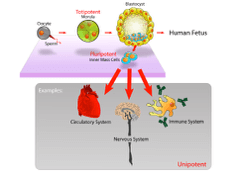
Cell division is required to produce organs After fertilisation, most cell differentiate from
pluripotent stem cells that come from the inner mass of the blastocyst After differentiation into
their specific cell type, the cells become unipotent
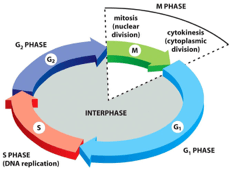
Stages of the cell cycle M phase (mitotic segregation) In this stage, the cell undergoes mitosis
(nuclear division) and undergoes cytokinesis (cytoplasmic division) G1 phase The first gap phase
can be divided into an early and a late stage, which is separated by restriction point Early G1
stage is mitogen-dependent and requires extrinsic growth factors or hormones (mitogens) which provide
the stimulatory signal to proceed forward The R point is kind of point of no return After this
point, the cell is committed to progressing to the next phase Hyperphosphorylation of Rb by
CDK4/cyclin D complex is important in passing through the R point (as hyperphosphorylation does not
inhibit e2F) Proteins called cyclins and cyclin-dependent kinases (CDKs) control progression in the
cell cycle by phosphorylation of regulatory proteins An example is the Rb (retinoblastoma tumour
suppressor protein) Unphosphorylated Rb binds to and inhibits e2F, the activation of which will
drive gene transcription and cause progression to late G1 CDK4 binds with cyclin D to phosphorylate
Rb which allows progression through the R point The point of this stage is for the cell to grow in
size and to synthesise RNAs and proteins required for DNA synthesis (replication) S phase Active
replication of chromosomes and centromeres takes place (DNA replication) G2 phase Phase that
precedes mitosis Cells increase in size further -> DNA synthesis completion is checked G0
phase Phase that non-dividing cells are at; also called quiescence
Alternating and completing S and M phases sequentially is critical to ensure maintenance of ploidy
Gap phases (G1 and G2) separate S and M phase in somatic cell cycles
Dysregulation of the cell cycle in cancer
Hallmarks of cancer cells Growth signal autonomy Resistance to inhibitory growth signals
Resistance to apoptosis Unlimited replicative capacity These first four relate to cell cycle and
cell death deregulation Sustained angiogenesis Tissue invasion and metastasis Apoptosis
One of the most noticeable features of apoptosis is condensation of the nucleus and its fragmentation
into smaller pieces, a highly distinctive event that is not seen under any other circumstances
Nuclear DNA is hydrolysed into numerous fragments, often in mutliples of 200bp The Golgi, ER and
mitochondrial networks also undergo pronounced fragmentation during apoptosis, and numerous proteins are
released from the mitochondrial intermembrane space Mainly, cytochrome c The release of
cytochrome c drives the assembly of a caspase-activating complex that contains caspase 9 and Apaf-1
(called the apoptosome) on release into the cytosol Hundreds of proteins undergo restricted
proteolysis during apoptosis mainly by caspases Thus, in mammals, the apoptotic signalling was found
to be:
Dysregulation of cell death in cancer Apoptotic cell clearance The terminal event of the
caspase-mediated demolition phase of apoptosis and removal and consumption of the dead cell by
phagocytes This is a critical step in the pathway as it means that dead cells are removed with their
plasma membranes intact The generation of signals such as ‘eat me’ and binding sites for phagocytes
and the release of chemoattractant molecules represent the last acts of the dying cell Phagocytes
are equipped with receptors that specifically detect engulfment signals on the apoptotic cell Every
cell in a multicellular organism has the potential to die by apoptosis However, tumour cells often
have faulty apoptotic pathways These defects not only increase tumour mass, but also render the
tumour resistant to therapy P53 is a key player in apoptosis induction in tumour cells P53 is
inhibited by MDM2, a ubiquitin ligase, that targets p53 for destruction by the proteasome MDM2 is
inactivated by binding to ARF Cellular stress, including that induced by chemotherapy or irradiation
activates p53 either directly, by inhibition of MDM2, or indirectly by activation of ARF (ARF binds to
MDM2 and removes it thus relieves p53 from inhibition) ARF can also be produced by proliferative
oncogenes such as Ras Active p53 transactivates pro-apoptotic genes such as Bax, Noxa, CD95 and
Trail-R1 to promote apoptosis
The Bcl-2 family of proteins This family of proteins are critical regulators of apoptosis through
their ability to control cytochrome c release from mitochondria The Bcl2 family is divided into
three groups based on their BCL-2 homology (BH) domain organisation Anti-apoptotic Bcl2 proteins
Bcl2 and its close relatives Bcl-XL, MCL1, BCL2a1 and Bcl-W all have BCL-2 homology (BH) domains and all
block apoptosis They do that by preventing BH3-only protein-induced oligomerisation of the
pro-apoptotic Bcl2 family members Pro-apoptotic Bcl2 proteins These are proteins such as BAX
and/or BAK in mitochondrial outer membranes Their protein-induced oligomerisation would lead to the
efflux of cytochrome c The anti-apoptotic Bcl2 proteins differentially bind to the BH3-only proteins
BID and BIM from the pro-apoptotic BH3-only group interact with all anti-apoptotic Bcl2 proteins,
whereas NOXA and PUMA interact with only certain anti-apoptotic members BH3-only proteins
Comprise 8 members (BID, BAD, BIM, BIK, BMF, NOXA, PUMA and HRK), all of which promote apoptosis when
overexpressed These share only a little sequence homology apart from the BH3 motif and they are
regulated in distinct ways NOXA and PUMA are up-regulated by p53 BID is activated through
proteolysis by caspase-8 to generate tBID
Pro-survival (Anti-apoptotic) Bcl2 proteins and pro-apoptotic proteins (such as Bax or Bak) control pore
formation Pore formation and leakage of cytochrome c results in loss of mitochondrial outer membrane
potential (MOMP) Bcl2 proteins are over-expressed in various types of non-Hodgkin’s lymphoma
Apoptosis inhibition via this pathway, or others, is a feature of cancerous cells Elevated levels of
Bcl-2 allow cells to ignore signals to undergo apoptosis Thus, Bcl2 is an oncogene The Bcl-2
gene has been identified as a gene directly involved in the consistent chromosome translocation t(14;18)
found in approximately 90% of human follicular lymphoma cases The translocation of the Bcl2 gene on
the chromosome band 18q21.3 results in consistent expression of the Bcl2 protein This Bcl2 fusion is
an oncogene and plays a crucial role in follicular lymphomagenesis
Hints & Tips on how to study cell cycle and apoptosis
The study of cell cycle can be rather daunting for students, as it appears to involve many genes
implicated in its regulation. The key to the cell cycle is to understand the role of cyclins,
cyclin-dependent kinases (CDKs) and the different ‘checkpoints’.
Cell cycle can be thought of just like any process. Take for example the manufacturing of a single coke
bottle: a highly automated process that has a few ‘checkpoints’ to make sure that the manufacturing line
is progressing as expected. This is exactly what happens during the cell cycle; however, this time, the
‘checkpoints’ are biochemical i.e. made up of proteins and enzymes that co-ordinate the process in a
complex fashion. For example, the key player in the initiation of the G1 phase is the checkpoint
controlled by the Rb protein that keeps the elongation factor E2F in check, thus preventing progression
in the cycle. Only when there is enough Cyclin D to bind the kinases CDK4 or CDK6, the kinases become
active and hyperphosphorylate Rb, thus removing the inhibition on E2F, allowing the progression of the
cycle past this checkpoint. Other signalling molecules control cyclin D, thus providing various levels
of cell cycle control.
Indeed, any student who is studying cell cycle should be focussing mainly on these checkpoints: the R
point, the G1/S checkpoint and the G2/M checkpoint and know what Cyclins and CDKs are associated with
these. Knowing these molecules will enable the student to then recognise what will happen should any of
these proteins become dysregulated. For example, it is know clear that if there are mutations in the Rb
gene, there will be an inadequate inhibition at the R point and the cell cycle will thus be overactive.
In patients who have Rb mutations, retinoblastomas i.e. cancers of the retina, occur.
Every cell has a limit of divisions it can perform – its so-called Hayflick limit. This limit is
determined by telomere shortening with every cell division. Once telomere shortens beyond that limit,
the cell undergoes programmed cell death – apoptosis in order to prevent the increased risk of any
genetic mutations. Apoptosis is therefore a key mechanism that our cells use to control the
three-dimensional architecture of tissues. It is therefore not surprising that any dysregulation in
apoptosis, or its evasion, may lead to tumour growth. For this reason, it is important to know the key
players in this process: the pro- and anti-apoptotic proteins our cells encode to strike the right
balance between survival and death, in both cases with the aim of providing a beneficial outcome to
tissue architecture. Cell stress therefore induces pro-apoptotic proteins as many mechanisms within the
cell become dysregulated, and thus their potential to grow out of control increases. The Bcl-2 family of
proteins are the key to apoptosis, and need to be known by any student attempting to understand this
process. In order to facilitate the best understanding of this, it is advisable to separate this family
into groups of pro- and anti-apoptotic proteins and know how they are linked to those molecules involved
in the cell cycle.
Cancer biochemistry is indeed a complex subject and does require memorisation of rather difficult names
for one to do well in exams. However, I believe the important message here is to practice layered
learning: learn the concepts first and then attach names to outlined processes. My students also
frequently benefit from interactive methods, such as online quizzes, where I test their knowledge in
simple MCQ, or extended-matching question formats.
We have hundreds of tutors available right now to help you improve and succeed. From a
one hour session online to a full academic year of face to face lessons, all it takes is five minutes for us
to take down your information. We can then find you the most suitable tutors.